
Introduction
Reproductive efficiency is declining on U.S. dairy herds. There are numerous factors that affect reproductive efficiency on individual farms. These include heat detection rate, heat stress, semen quality, bull fertility, AI technique, nutrition, body condition score, clinical and subclinical mastitis, cow comfort, and many other factors related to whole farm management, individual farm managers and laborers, and individual cows. This manuscript will briefly summarize two recent research areas. The first section of the manuscript discusses a recent study on body condition score. This study was part of a larger study that evaluated numerous factors simultaneously to determine the management factors most related to reproductive performance (Caraviello et al., 2006). This study used a machine learning technique to simultaneously evaluate numerous factors (341 different variables) that may be affecting reproductive efficiency on U.S. dairies. Interestingly, one of the top factors that was found to be closely associated with reproductive performance was the amount of bunk space per cow in the breeding pen. The predicted percentage of cows that were pregnant by 150 days tended to increase linearly as bunk space increased from 30 to 60 cm (from 35% to 70%). Thus, overcrowded cows (<36 cm of bunk space/cow) had a much lower percentage of pregnancy at 150 days in milk than cows with more bunk space.
Another factor that was closely related to reproductive efficiency was body condition score. This part of the study will be discussed in more depth in Section II below. In addition, milk production was found to be associated with reproductive efficiency in a fairly complex manner. Section III of this manuscript is based on recent research in our laboratory that has found an intriguing biological link between high feed consumption and reproductive efficiency caused by elevated metabolism of hormones. While this elevated hormone metabolism does not explain all aspects of nutritional effects on reproduction in dairy cows, it is becoming clear that hormone metabolism is a key missing link between high milk production and certain changes in reproductive efficiency such as decreased expression of estrus.
Please check this link first if you are interested in organic or specialty dairy production.
Body Condition Score and Reproductive Performance
There is substantial evidence linking body condition score (BCS) to reproductive performance in dairy cows. Pryce et al. (2000), evaluating records from 44,674 first lactation cows, found that BCS at 1 mo after parturition had estimated genetic correlations with calving interval of -0.40 and -0.22, before and after adjusting for milk yield, respectively. They also reported that, although the change in BCS between 1 mo and 4 mo postpartum had a high genetic correlation with fertility, the average or absolute BCS for a given cow had an even stronger relationship with fertility. These authors indicated that a single BCS measured early in lactation could be used as an indirect selection criterion for fertility. Other authors have also reported a genetic relationship between BCS and fertility (Dechow et al., 2004; Pryce et al., 2001).
The negative impact of low BCS on female fertility could be due to a number of factors, including a longer time from parturition to onset of ovarian activity among cows with low BCS. Veerkamp et al. (2000) used a milk progesterone assay to indicate the onset of luteal activity and obtained an estimated genetic correlation of -0.54 between onset of luteal activity and body weight (which is highly correlated with BCS) at 100 d postpartum. The same study reported an estimated genetic correlation of -0.80 between onset of luteal activity and body weight change from calving to 100 d postpartum. Royal et al. (2002) later reported an estimated genetic correlation of -0.84 between commencement of luteal activity and BCS.
It appears that BCS measured 30 to 60 d postpartum has the strongest relationship with reproductive performance and that multiple BCS measurements can increase this relationship (Dechow et al., 2001; Pryce et al., 2000). Nevertheless, it may be impractical to measure BCS repeatedly at specific stages of lactation for every cow in a herd. Therefore, comparing BCS collected at different points during the lactation to predetermined standards or thresholds may provide valuable, cost-effective information for herd managers and dairy consultants. Therefore, the objectives of this study were two-fold. First, to develop a BCS fault system that can predict reproductive performance both at the herd level and at the individual cow level. Second, to compare the predictive ability of BCS faults (or lack thereof) on the day of insemination with BCS treated as a continuous variable.
We used reproductive data from 153 large commercial dairies that participate in the Alta Advantage progeny-testing program. After editing, the data set had 17,587 lactation records from 9,516 cows that calved from 2000 to 2004 on 153 large (average of 613 ± 46 lactating cows) commercial dairies across the United States. We obtained BCS on a subset of these cows and did an initial analysis to find the average values of BCS for cows at different stages of lactation (shown in Figures 1, 2, and 3). This was done to determine the normal BCS in these herds and to set a “normal” value for all stages of lactation that could be used to determine the “BCS status” of each herd. Mean BCS for dry cows was 3.22; this declined to 2.88 by 30 d postpartum and declined further to 2.70 by 50 d postpartum. Mean BCS subsequently increased to 2.75 by 80 d postpartum and stabilized at around 2.85 by 200 d postpartum. Figure 1 shows the distribution of BCS values for a herd with few BCS “faults.” Most cows in this herd fell above the polynomial curve, and the percentage of BCS faults was only 2%. Figure 2 shows an example herd with substantial variation in BCS, such that many cows fall well above or well below the polynomial curve. The overall percentage of BCS faults in this herd was 15%. Lastly, Figure 3 shows a herd with very poor BCS; few cows in this herd are above the polynomial curve, and the overall fault rate in this herd was 54%.

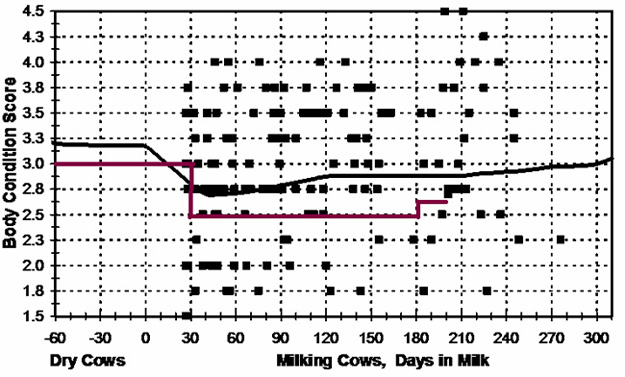

The herds in this study ranged from nearly 0% to more than 60% BCS faults. The relationship between percentage of BCS faults and reproductive performance was evaluated at the herd level by comparing herd BCS status with the percentage of cows pregnant by 150 d postpartum. Pregnancy status at 150 d postpartum was chosen as the dependent variable because, as discussed above, it is a robust measure of overall herd reproductive performance. In addition to the herd level analysis, we also evaluated BCS status at the individual cow level by comparing pregnancy status at 150 d and conception rate at first service with the presence or absence of a BCS fault for an individual cow.
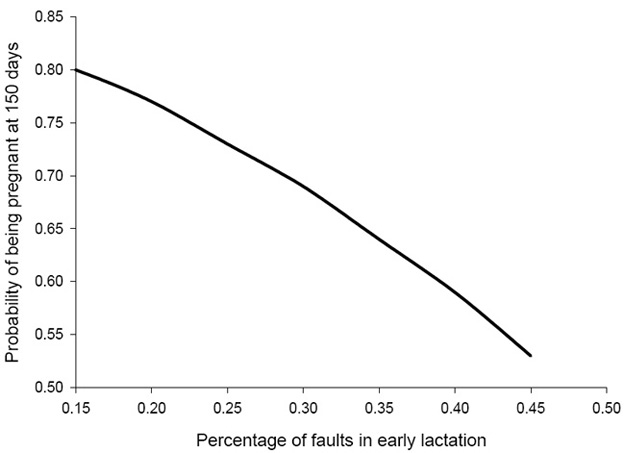
Application of a logistic regression model at the herd level showed that the proportion of cows pregnant by 150 d postpartum decreased linearly as the percentage of BCS faults increased (Figure 4). The probability of pregnancy by 150 d postpartum was 0.80 for herds with 15% BCS faults, as compared with 0.53 for herds with >45% BCS faults.
Cow level evaluation of the relationship between BCS faults and probability of pregnancy by 150 d postpartum confirmed trends that were observed at the herd level. Individual cows with BCS faults had significantly (P < 0.05) lower probability of becoming pregnant by 150 d postpartum (0.61) than cows without faults (0.74).
Thus, we have developed a simple system to separate herds according to BCS status determined at a single farm visit. This BCS “fault” system can be used to identify herds in which BCS problems may be the cause of poor overall reproductive performance. Alternatively, herds can be identified in which BCS faults should not be viewed as a likely cause of poor reproductive performance, and therefore other areas should be examined.
High Feed Consumption and Reproductive Efficiency Are Linked by High Hormone Metabolism
This section of the manuscript will not attempt to review all of the nutritional factors that can potentially alter reproductive efficiency in dairy cattle. This has been reviewed in numerous other publications and would require a complete volume for adequate review. We will focus on a critical link that has become increasingly relevant to our understanding of nutrition and reproduction. This link is very different from the link discussed above concerning BCS and reproduction. Negative energy balance and the resulting low BCS cause a delay in the time to first ovulation and thus increase the percentage of cows that are anovular (not ovulating). The increased percentage of anovular cows is not clearly tied to milk production in most studies. However, there are other key reproductive efficiency measures that are clearly related to milk production. This review will focus on two key reproductive measures, efficiency of heat detection and double ovulation rate. These are chosen because of the clear relationship to milk production. We will use these measures to build a model for how reproductive efficiency can be dramatically altered by the high feed consumption associated with high milk production. These two measures are used to clearly demonstrate the physiological model that can explain the relationship between high feed consumption and decreased reproductive efficiency. However, this model becomes much more complicated when trying to explain a much more complex reproductive variable such as conception rate. Nevertheless, it seems likely that this model will also be important in explaining some aspects of this variable as well.
Duration of Estrus
It is clear that low rates of estrous detection are reducing reproductive efficiency on commercial dairy farms. Indeed, Washburn et al. (2002) reported a decrease from 50.9% in 1985 to 41.5% in 1999 for estrous detection rates in Holstein dairy herds in the southeastern United States. However, studies have reported both a negative relationship between level of milk production (Harrison et al., 1990; Harrison et al., 1989) or no relationship (Fonseca et al., 1983; VanEerdenburg et al., 2002) using visual observation twice daily to measure expression of estrus. We have recently completed a study in which we evaluated the duration of estrus in a group of lactating dairy cows using the HeatWatch system (Lopez et al., 2004). This system allowed continuous monitoring of all mounts 24 h per day and can be used to calculate the duration of estrus in individual dairy cows. Cows with milk production above the herd average (~ 40 kg/d) had shorter (P < 0.001) duration of estrus (6.2 ± 0.5 h) than cows with lower milk production (10.9 ± 0.7 h). This effect was not due to a parity effect because separate analysis of primiparous and multiparous cows showed a similar effect. Figure 5 shows the relationship between level of milk production and duration of estrus. In order to consistently observe this strong negative relationship between level of milk production and duration of estrus, it is critical that milk production data be collected close to the time of estrus, that only data from ovulations after the first postpartum ovulation be utilized (first ovulation has low expression of estrus), that all ovulations be consistently monitored throughout the observation period (to avoid false estrus or missing data from ovulations), and that duration of estrus be monitored on a continuous basis with an electronic heat monitoring system.

As discussed below, we theorize that high milk production leads to decreased circulating estradiol concentrations producing decreased duration of estrus. Decreased estradiol could also cause increased follicular size by delaying the time to estradiol-induction of estrus, GnRH/LH surge, and ovulation in high-producing cows.
Double Ovulation Rate
Another reproductive trait that has been directly linked to milk production is double ovulation rate (for a more complete review, see (Lopez et al., 2005; Wiltbank et al., 2000). From a practical standpoint, double ovulation rate appears to be the underlying cause of increased twinning rate in lactating dairy cows, with 93% of twins being nonidentical (Silva Del Rio et al., 2004). Numerous factors have been recognized as possible regulators of twinning rates, including age of dam, season, genetics, use of reproductive hormones or antibiotics, ovarian cysts, days open, and peak milk production (reviewed in Wiltbank et al., 2002). In a large study on risk factors for twinning, Kinsel et al. (1998) concluded, “the single largest contributor (> 50%) to the recent increase in the rate of twinning is the increase in peak milk production.” We performed a study in which we evaluated double ovulation rate in 240 dairy cows (Fricke et al., 1999) that had ovulation synchronized with the Ovsynch protocol. Ovulation was determined by transrectal ultrasonography at the time of the second GnRH injection and 48 h later. The mean milk production, determined 3 d before ovulation, was 40.7 ± 0.8 kg/d, and cows were segregated by whether they were below or above the mean value. Double ovulation rate in cows that were above average production was 20.2% compared to 6.9% in those below average (P < 0.05). This difference was similar regardless of lactation number. Recently, we reported results of a study (Lopez et al., 2005) that evaluated naturally ovulating dairy cattle and found a similar relationship between milk production and double ovulation rate (Figure 6). Cows that produced less than 40 kg/d had a very low double ovulation rate, whereas cows producing above 50 kg/d had more than a 50% double ovulation rate. It is surprising that there is such a dramatic inflection point in double ovulation rate as milk production increases above 40 kg/d, and it is still unclear what physiological changes occur as milk production increases above this critical value. This increase in double ovulation rate is likely to continue to increase twinning rate in dairy herds as milk production increases. It is also clear that this effect of milk production is most related to the level of production within the 2 wk before the cow ovulates and not to total milk production during the entire lactation. This effect was also similar when a more extensive regression model was used for analysis and when multiparous and primiparous cows were analyzed separately (Lopez et al., 2005). As with duration of estrus, the first postpartum ovulation differed from other ovulations with a high double ovulation rate that was unrelated to milk production.

How Does High Milk Production Change Duration of Estrus or Double Ovulation Rate?
We have synthesized a good deal of information into a model that we think helps explain some of the changes in reproduction due to high milk production (Figure 7). Lactating cows have greater energy requirements than non-lactating cows (for example, a cow producing 50 kg/d of milk will require 53 Mcal/d of energy versus 12.5 Mcal/d for a non-lactating cow; NRC, 2001). The high feed consumption required to meet these energy requirements leads to a dramatic increase in blood flowing to the digestive tract to pick up these nutrients. All blood that flows through the digestive tract has to flow through the liver. Therefore, there will be a very high amount of blood flowing to the liver in cows with high feed consumption (such as high-producing dairy cows). This may seem trivial, but the liver is the organ that rids the body of many compounds including hormones like progesterone and estrogen. Therefore, high feed consumption, through this simple pathway, will lead to a very high amount of the estrogen and progesterone in the body being metabolized (broken down) in the liver. This would cause lower estrogen and progesterone levels in the blood, and this could cause problems with many different aspects of reproduction. For example, low estrogen would cause the animal not to come into heat or to be in heat only for a very short period of time. Thus, this simple model could potentially explain some of the results described in the sections above.

Practical Implications for Reproductive Management
The next section will briefly suggest some practical implications and reproductive management strategies for each of these areas.
1. Decreased Duration of Estrus Due to High Milk Production:
What does this practically mean for a dairy farm? We used the data on duration of estrus versus milk production to analyze what would happen to heat detection efficiency for cows with different levels of milk production. Figure 8 shows the probability of detecting a cow in heat with different frequency of heat detection. If a cow is producing about 70 lb of milk per day, a four-time per day heat detection program will detect about 90% of cows that are in estrus. However, this same program (four times/d) will only detect about 50% of cows in heat if they are producing above 100 lb/d. This result gets even worse if heat detection is done only twice per day or once per day. It should be noted that all of the probabilities in this analysis were based on actual ovulation by the cows (detected by ultrasound). Although some producers will say that the high-producing cows are not cycling, they are cycling normally. However, producers do not detect the cows in heat because of the short period in which they are in heat. Increasing number of times that cows are checked for heat can help to solve this problem. Many producers are using heat detection aids such as tail chalk to help find cows that are in showing heat at a time that they are not present. This can be critical because high-producing cows are showing heat for only 4 hours or less in many cases. Most dairy producers in the United States are incorporating timed AI programs, such as Ovsynch, into their reproductive management programs to allow high-producing cows to be bred in a timely manner.

2. Treating Anovular Cows:
Although level of milk production is not normally associated with incidence of anovulation, dairy producers still need to design programs to treat anovular cows. Generally, 20% of dairy cows will not be cycling by 70 days after calving. This percentage will increase if there are a high percentage of cows with low body condition scores (2.5 or less). These cows need to be quickly assigned to a hormonal program (and possibly nutritional program if they have low body condition score) that will start the cows cycling. An Ovsynch program alone is not the ideal treatment for anovular dairy cows. Use of a CIDR or pretreatments with GnRH should be incorporated into these programs to be optimal treatments for non-cycling dairy cows.
3. Increasing Double Ovulation Rate (and Twinning Rate) with Increasing Milk Production:
From a practical standpoint, it appears that there may be little that we can do to change this trend. Using Ovsynch does not seem to increase or decrease double ovulation with double ovulation related to milk production whether we look after a hormonal synchronization program or a natural estrus. Obviously, not all double ovulations result in twins, but increasing double ovulation rate will almost surely result in increased twinning rates on higher-producing farms. It seems clear that the main increase occurs after cows are producing about 90 lb/d. Thus, we must anticipate that we will have a dramatic increase in double ovulation rate in cows producing over 90 lb/d, and this will result in an increase in twinning rate in cows that conceive during this time of high milk production. We must align our management procedures to deal with this increasing twinning rate if we are increasing milk production into this range in our dairies. First, we must set a program to diagnose twins. Second, we should set up procedures to manage cows that are likely to have twin births. Twinning cows will calve earlier (10 to 14 d on average) and are likely to have more problems during the calving process. These twin calving cows were, on average, our highest-producing cows during the previous lactation; therefore, we must carefully design our calving and early lactation procedures with these twinning cows in mind.
4. Decreasing Conception Rate Due to Higher Milk Production:
As mentioned above, there are many different factors that impact conception rate in lactating dairy cows, and higher milk production is just one of these factors — and in many farms, it may be a fairly minor factor. The effect of production on fertility is dramatically amplified during hotter times of the year. This is because there is a greater increase in body temperature as cows increase milk production. This increase in body temperature leads to decreased reproductive success, particularly death of the early embryo.
From a practical viewpoint, we have tried to utilize the information that many of the problems with fertility in dairy cows appear to occur during the first week after breeding. We hypothesized that we could improve reproduction just by transferring a good-quality embryo at 7 days after expected time of AI. So in a fairly large experiment, we compared the conception rate in our herd when cows were bred either by AI or by embryo transfer (ET). Unfortunately, ET did not improve conception rates compared to AI. Therefore, it appears that there may be multiple problems with fertility in lactating dairy cows, and just transferring an embryo from a heifer does not appear to remove all of the fertility problems from the lactating cow. Many laboratories are currently experimenting with a number of changes in timed AI programs that may increase conception rates in high-producing dairy cows. There are numerous intriguing possibilities, but they still lack sufficient data to allow recommendation at this time. One of the simplest recommendations is to set up a proper time of AI, particularly during the Ovsynch program. Many producers are currently using a CoSynch program (AI at the same time as second GnRH injection), but we have found that this will result in a substantial decrease in conception rates. The simple strategy of optimizing the time of AI after the second GnRH injection of Ovsynch (16 h from GnRH to AI) should increase the number of pregnancies by ~33%. This is a substantial improvement (30% to 40%) and is probably the clearest and easiest management strategy that has been experimentally validated and can be immediately implemented to increase conception rates on dairy farms. Many other management strategies based on the physiological models developed above are still in the experimental stages.
Author Information
Milo C. Wiltbank
Kent A. Weigel
Daniel Z. Caraviello
University of Wisconsin
References
Caraviello, D.Z., K.A. Weigel, M. Craven, D. Gianola, N.B. Cook, K.V. Nordlund, P.M. Fricke, and M.C. Wiltbank. 2006. Analysis of reproductive performance of lactating cows on large dairy farms using machine learning algorithms. J. Dairy Sci. 89:4703-4722.
Dechow, C.D., G.W. Rogers, L. Klei, T.J. Lawlor, and P.M. VanRaden. 2004. Body condition scores and dairy form evaluations as indicators of days open in US Holsteins. J. Dairy Sci. 87:3534-3541.
Fonseca, F.A., J.H. Britt, B.T. McDaniel, J.C. Wilk, and A.H. Rakes. 1983. Reproductive traits of Holsteins and Jerseys. Effects of age, milk yield, and clinical abnormalities on involution of cervix and uterus, ovulation, estrous cycles, detection of estrus, conception rate, and days open. J. Dairy Sci. 66:1128-1147.
Fricke, P.M., M.C. Wiltbank. 1999. Effect of milk production on the incidence of double ovulation in dairy cows. Theriogenology 52:1133-1143.
Grummer, R.R., and D.J. Carroll. 1988. A review of lipoprotein cholesterol metabolism: Importance to ovarian function. J. Anim. Sci. 66:3160-3172.
Harrison, R.O., S.P. Ford, J.W. Young, A.J. Conley, and A.E. Freeman. 1990. Increased milk production versus reproductive and energy status of high producing dairy cows. J. Dairy Sci. 73:2749-2758.
Harrison, R.O., J.W. Young, A.E. Freeman, and S.P. Ford. 1989. Effects of lactational level on reactivation of ovarian function, and interval from parturition to first visual oestrus and conception in high-producing Holstein cows. Anim. Prod. 49:23-28.
Kinsel, M.L., W.E. Marsh, P.L. Ruegg, and W.G. Etherington. 1998. Risk factors for twinning in dairy cows. J. Dairy Sci. 81:989-993.
Lopez, H., D.Z. Caraviello, L.D. Satter, P.M. Fricke, and M.C. Wiltbank. 2005. Relationship between level of milk production and multiple ovulations in lactating dairy cows. J. Dairy Sci. 88:2783-2793.
Lopez, H., L.D. Satter, and M.C. Wiltbank. 2004. Relationship between level of milk production and estrous behavior of lactating dairy cows. Anim. Reprod. Sci. 81:209-223.
Pryce, J.E., M.P. Coffey, and G. Simm. 2001. The relationship between body condition score and reproductive performance. J. Dairy Sci. 84:1508-1515.
Pryce, J.E., M.P. Coffey, and S. Brotherstone. 2000. The genetic relationship between calving interval, condition score and linear tupe and management traits in pedigree registered Holstein dairy cows. J. Dairy Sci. 83:2664-2671.
Royal, M.D., J.E. Pryce, J.A. Woolliams, and A.P.F Flint. 2002. The genetic relationship between commencement of luteal activity and calving interval, body condition score, production, and linear type traits in Holstein-Friesian dairy cattle. J. Dairy Sci. 85:3071-3080.
Sangsritavong, S., D.K. Combs, R. Sartori, and M.C. Wiltbank. 2002. High feed intake increases blood flow and metabolism of progesterone and estradiol-17β in dairy cattle. J. Dairy Sci. 85:2831-2842.
Silva Del Rio, N., B.W. Kirkpatrick, and P.M. Fricke. 2004. Observed frequency of monozygotic twinning in lactating Holstein cows. J. Dairy Sci. 87 (Suppl. 1):65 [Abstract].
Van Eerdenburg, F.J.C.M., D. Karthaus, M.A.M. Taverne, I. Meries, and O. Szenci. 2002. The relationship between estrous behavioral score and time of ovulation in dairy cattle. J. Dairy Sci. 58:1150-1156.
Vasconcelos, J.L.M., S. Sangsritavong, S.J. Tsai, and M.C. Wiltbank. 2003. Acute reduction in serum progesterone concentrations after feed intake in dairy cows. Theriogenology 60:795-807.
Veerkamp, R.F., J.K. Oldenbroek, H.J. Van der Gaast, and J.H.J. Van der Werf. 2000. Genetic correlation between days until start of luteal activity and milk yield, energy balance, and live weights. J. Dairy Sci. 83:577-583.
Washburn, S.P. W.J. Silvia, C.H. Brown, B.T. McDaniel, and A.J. McAllister. 2002. Trends in reproductive performance in southeastern Holstein and Jersey DHI herds. J. Dairy Sci. 85:244-251.
Wiltbank, M.C., P.M. Fricke, S. Sangsritavong, R. Sartori, and O.J. Ginther. 2000. Mechanisms that prevent and produce double ovulations in dairy cattle. J. Dairy Sci. 83:2998-3007.