
Contents |
Abstract
Prospects for improving female fertility in dairy cattle via genetic selection are reviewed. Today’s high-producing cows have shorter estrous cycles, fewer standing events, shorter duration of estrus, and more frequent multiple ovulations. Although high milk production is often implicated as the cause of impaired fertility, the impact of inadequate body condition appears to be greater, as the latter has a significant impact on probability of conception, rate of embryonic loss, and proportion of anestrous animals. Genetic improvement of female fertility can be achieved by indirect selection for productive life (PL) or body condition score (BCS), or by direct selection for traits such as daughter pregnancy rate (DPR). Most leading dairy countries have implemented genetic evaluation systems for female fertility in the past decade, but refinement of these systems to account for hormonal synchronization, differences in the voluntary waiting period, exposure to natural service bulls, and other confounding factors is warranted. Recent work has focused on the development of data collection and genetic evaluation systems that will allow selection of bulls that have daughters that are resistant to common health disorders, including mastitis, lameness, ketosis, displaced abomasum, and metritis. Such systems will allow selection of animals that can remain healthy and fertile while producing large quantities of milk.
Introduction
The challenges associated with achieving pregnancy in modern, high-producing dairy cows have received considerable attention from scientists, veterinarians, and farmers in recent years. Today’s dairy cows tend to have lower conception rate, greater days open, and greater likelihood of culling due to infertility than their counterparts from two or three decades ago. Genetic selection programs have led to rapid gains in milk yield and conformation traits; but performance for traits such as female fertility, longevity, and susceptibility to disease has tended to decline. While it is impossible to completely disentangle the effects of selection from simultaneous changes in nutrition, cow care, and reproductive management, it is clear that geneticists failed to pay adequate attention to health, fertility, and longevity traits until the past decade. The magnitude of genetic variation in such traits is surprising, and we are now poised to take advantage of recent research and development efforts regarding the definition, measurement, and genetic analysis of these traits.
The objective of this paper is to review the relationships between female fertility and other economically important dairy traits and to discuss opportunities for improving reproductive performance through direct selection of highly fertile animals or indirect selection of animals that maintain adequate body condition and resist metabolic and infectious diseases during lactation.
Please check this link first if you are interested in organic or specialty dairy production
Relationships between Fertility and Other Economically Important Traits
Milk Yield and Fertility
Milk production of dairy cows on modern commercial farms has roughly doubled over the past four decades. First parity cows on large commercial dairy farms typically peak at 40 to 45 kg/d, while second and later parity cows typically peak at 50 to 55 kg/d. Furthermore, each group typically sustains daily milk production of 40 kg/d or more during the first seven months postpartum. Therefore, one might expect differences in the reproduction of high-producing cows, as compared with low-producing cows or yearling heifers.
Lopez et al. (2005) discussed some of the differences between the reproductive biology of lactating Holstein cows and yearling Holstein heifers. In particular, Lopez et al. (2005) noted that lactating cows have shorter duration of estrus (7 to 8 hr vs. 11 to 14 hr), longer and more variable estrous cycles (20 to 29 d vs. 20 to 23 d), larger diameter of ovulatory follicles (16 to18 mm vs. 14 to 16 mm), and greater rates of anovulation (20 to 30% vs. 1 to 2%), multiple ovulation (20 to 25% vs. 1 to 3%), and pregnancy loss (20 to 30% vs. 3 to 5%).
Lopez et al. (2005) also documented differences in these characteristics between lactating cows according to levels of milk production. They (Lopez et al., 2005) used the HeatWatch system (DDx Inc., Denver, Colorado) to monitor the estrous characteristics of 146 high-producing Holstein cows (46.4 kg/d for the 10 d preceding estrus) and 177 low-producing Holstein cows (33.5 kg/d for the 10 d preceding estrus). High-producing cows had shorter duration of estrus (6.2 hr vs. 10.9 hr), fewer standing events (6.3 vs. 8.8), and shorter standing time per event (21.7 sec vs. 28.2 sec). Duration of estrus decreased linearly from 14.7 hr for cows milking 25 to 30 kg/d to 2.8 hr for cows milking 50 to 55 kg/d. In addition, the percentage of cows with multiple ovulations increased from 0.0% for cows milking between 25 and 30 kg/d to 51.6% for cows between 50 and 55 kg/d.
The rate of early embryonic loss in Holstein cows is also a major concern, as noted in several recent studies that have used ultrasound for pregnancy detection at 27 to 31 d after breeding, followed by pregnancy confirmation via rectal palpation at 39 to 48 d after breeding. Reported rates of embryonic loss during this interval ranged from 0.70 to 1.40% per day (e.g., Cartmill et al., 2001; Cerri et al., 2004; Santos et al., 2004). However, estimates of the rate of embryonic loss (particularly those from commercial farms) may be biased upward by false positive diagnoses at the early ultrasound exam, as most veterinarians tend to use caution when declaring cows as non-pregnant in herds that use hormonal resynchronization programs.
On large western dairy farms, mean veterinary-confirmed conception rates of Holstein cows at 75 d after breeding were nearly constant over the first five inseminations (0.30, 0.31, 0.31, 0.29, and 0.28, respectively), while means for Jersey cows declined linearly from the first through fifth insemination (0.42, 0.38, 0.34, 0.29, and 0.27, respectively). Mean conception rate at first service tended to decline with age in both breeds (0.35, 0.29, 0.28, 0.26, and 0.25, respectively, for first through fifth parity Holsteins and 0.44, 0.43, 0.41, 0.39, and 0.37, respectively, for first through fifth parity Jerseys), though the rate of decline was less noticeable for repeat inseminations than for first insemination (Weigel, 2006 (unpublished)). Both breeds have been selected for many generations under similar management conditions, and both have made rapid genetic progress over the past three decades (mean mature equivalent 305 d milk yield increased from 6,904 to 11,608 kg in Holsteins and from 4,461 kg to 8,273 kg in Jerseys from 1970 to 2000). Differences in mean conception rate within the Holstein breed were found among cows at different levels of daily milk yield, but such differences were smaller than one might expect (Weigel, 2005 (unpublished)). Mean conception rates at 75 d after breeding were 0.33, 0.33, and 0.32 for primiparous Holstein cows that averaged < 27 kg/d, 27 to 36 kg/d, and > 36 kg/d, respectively, during the first 3 mo of lactation; whereas corresponding means were 0.28, 0.28, and 0.27 for multiparous Holstein cows that averaged < 36 kg/d, 36 to 45 kg/d, and > 45 kg/d, respectively. In Wisconsin Holsteins, Lopez et al. (2005) found no relationship between the percentage of cows exhibiting anovulatory condition and level of daily milk yield. The percentage of anovular cows was 27.8% for cows that were milking 25 to 30 kg/d and 26.3% for cows that were milking 50 to 55 kg/d (means for 5-kg intervals in between ranged from 21.7 % to 35.1%, with no apparent trend). In California Holsteins, Santos et al. (2004) found a weak, nonsignificant relationship between milk yield and rate of embryonic loss between 31 and 45 d after breeding, with rates of 9.7% for cows that were milking 36 kg/d and 12.7% for cows that were milking 52 kg/d. Thus, it does not appear that increased milk yield is solely responsible for the decline in mean reproductive performance.
Body Condition Score and Fertility
High milk production, whether achieved through genetic selection, enhanced nutrition, or improved management, is often implicated as the cause of health, fertility, and culling problems on modern dairy farms. However, a complex relationship exists between milk yield, health, and reproductive performance. High-producing cows tend to be more susceptible to metabolic disorders and infectious diseases, and these can lead to impaired fertility. On the other hand, healthy cows tend to have higher milk production and greater reproductive performance than unhealthy cows. Conversely, cows that remain nonpregnant for much of the lactation tend to achieve higher levels of total production because fewer resources are allocated to the developing calf. Thus, one must be cautious when attempting to formulate cause-effect relationships between these traits.
Many recent studies of the relationships between genetic merit, nutritional status, and reproductive performance have identified a common culprit – inadequate body condition. Modern dairy cows undergo a process of building tissue reserves in late lactation and during the dry period, depleting these reserves after calving and in early lactation, and rebuilding them again in late lactation. Inadequate body condition can lead to numerous problems in the lactating dairy cow, including greater susceptibility to metabolic disorders (e.g., ketosis or displaced abomasum) and impaired fertility. Several authors have noted that cows with poor body condition in early lactation or a significant loss in body condition during the lactation tend to have impaired reproductive performance, including increased time from parturition to onset of ovarian activity, lower conception rate, and increased days open (Pryce et al., 2000; Royal et al., 2002).
Lopez et al. (2005) reported an extremely strong relationship between anovulatory condition and body condition score (BCS). Among lactating U.S. Holstein cows that had a BCS ≤ 2.50 (on a 5-point scale), 83% were anovular. Corresponding percentages of anovular animals were 38% for cows with a score of 2.75; 34% for cows with a score of 3.00; 22% for cows with a score of 3.25, and 9% for cows with a score ≥ 3.50. In a large, well-managed Holstein herd in Wisconsin, Caraviello et al. (2006) reported a conception rate of 31% for cows with a BCS at breeding of 2.25; 36% for cows with a score of 2.50; 40% for cows with a score of 2.75, and 42% for cows with a score of 3.00 or 3.25.
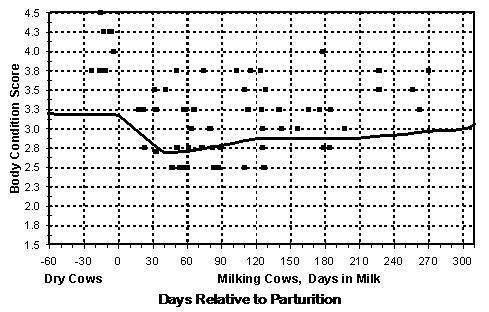
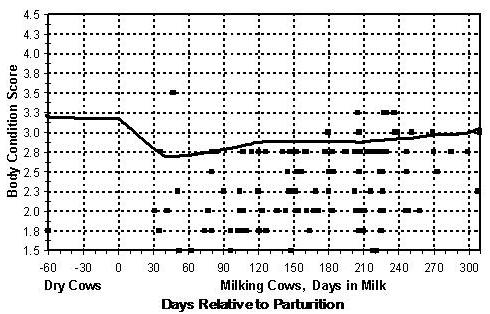
Caraviello et al. (2006) developed a system for evaluating the percentage of BCS “faults” on a commercial dairy farm. A single trained evaluator measured the condition score of 8,036 cows during single visits to 63 large dairy farms over a three-month period. Thresholds (i.e., minimum scores) for assigning a fault were 3.00 (on a 5-point scale) for cows that were 60 d prepartum to 30 d postpartum; 2.50 for cows that were 30 d postpartum to 180 d postpartum, and 2.75 for cows that were > 180 d postpartum (but not yet 60 d prepartum in the subsequent lactation). Figures 1A and 1B show herds with low and high percentages of BCS faults, respectively. The primary advantage of the fault system is that it provides a snapshot of the body condition status of a dairy herd in a single visit.
Although simple, the aforementioned fault system can be a powerful predictor of dairy cow fertility. Caraviello et al. (2006) evaluated the relationship between the percentage of BCS faults within a herd and the percentage of cows that became pregnant by 150 d postpartum. As the percentage of faults increased from 15% to 45%, the percentage of cows in the herd that were pregnant by 150 d postpartum decreased linearly, from 80% to 52%.
Because of recent concerns about the negative consequences of inadequate body condition in high-producing cows, the Holstein Association USA introduced a negative economic weight for dairy form, a measure of angularity that has an estimated genetic correlation of -0.80 with BCS (Dechow et al., 2004) into its selection index (commonly known at TPI) in 2005. It appears that historical selection pressure for cows that exhibited a high degree of “dairy character” or “angularity” (or perhaps cows that were simply too thin) may have been more harmful than selection for high milk production alone because antagonistic genetic correlations between fertility and BCS are greater than those between fertility and milk yield.
Mastitis and Fertility
During the past five years, several studies have considered the relationship between fertility and other health traits, such as mastitis. Kadarmideen et al. (2000) reported estimated genetic correlations between clinical mastitis and calving interval, services per conception, conception rate, and days to first service of 0.28, 0.41, -0.21, and 0.32; all of these relationships suggest that cows with more mastitis tend to have poorer fertility. Pryce et al. (1999) reported estimated genetic correlations of -0.58 between clinical mastitis and conception rate and 0.29 between clinical mastitis and calving interval. Likewise, the estimated genetic correlation between somatic cell score (SCS) and conception rate was -0.40, and the estimated correlation between SCS and calving interval was 0.14. Thus, it appears that cows with mastitis tend to have lower conception rate and longer calving interval.
Genetic Selection for Female Fertility
Genetic selection for health, fertility, and longevity has been practiced for more than two decades in some Scandinavian countries, and this has tended to stabilize (or at least lessen) the decline in mean performance for such traits. However, most other leading dairy countries did not pay serious attention to health, fertility, longevity until the mid-1990s. In the United States, the introduction of genetic evaluations for length of productive life (PL) in 1994 provided the first opportunity to improve female fertility through genetic selection (VanRaden and Klaaskate, 1993). Although length of PL is an indirect predictor of fertility, a limit of 10 months of PL per lactation was applied to ensure that cows with a regular calving interval would be favored. As shown in Figure 2, the genetic decline in daughter pregnancy rate (DPR), the measure of female fertility used for genetic evaluation of U.S. dairy sires, tended to stabilize around the time at which sire evaluations for length of PL became available.
Although indirect selection for improved fertility based on genetic evaluations for length of PL can be helpful, direct selection for improved fertility is more desirable. In the United States, genetic evaluations for DPR were introduced in 2003 (VanRaden et al., 2004). Input data are days open, computed from reported breeding dates for current cows and from calving interval for historical cows, and these are subsequently transformed to 21-day pregnancy rates for the purpose of genetic evaluation. The latter is a common, timely measure of reproductive efficiency on commercial dairy farms in the United States, and it is calculated as: (number of cows that became pregnant during a given 21-day period)/number of cows that were eligible for breeding at the beginning of the period). The pool of eligible cows includes those that have passed the voluntary waiting period but are not yet pregnant. Although 21-day pregnancy rate is typically used as a “herd measure” of reproductive performance, sire evaluations for this trait can be interpreted as the expected difference in 21-day pregnancy rate between the progeny groups of different sires (the 21-day pregnancy rate observation for an individual daughter can also be thought of as the number of 21-day opportunity periods required to achieve pregnancy). Differences between sires are dramatic – the highest and lowest available Holstein sires differ by 7.2% in DPR. Because a 1% difference in pregnancy rate corresponds to approximately four days open (VanRaden et al., 2004), daughters of the highest and lowest sires differ by roughly 29 days open per lactation. Mean DPR between sires in the top and bottom deciles is 4.9%, which corresponds to a difference of 20 days open per lactation. Breed differences also exist, as mean DPR for the Jersey breed is 4.6% higher than that of the Holstein or Brown Swiss breed.

DPR has been incorporated into all major selection indices used by U.S. dairy farmers, with a relative weight of 5 to 7% of the total economic value, so genetic improvement (or at least stabilization) of female fertility is expected. Furthermore, the estimated genetic correlation of -0.31 between milk yield and female fertility [Weigel, 2003 (unpublished)], although antagonistic, is small enough in magnitude to ensure the availability of some sires with high-producing and highly fertile daughters. Additional improvements will be made in genetic evaluations for DPR in the future, as the U.S. Department of Agriculture’s Animal Improvement Programs Laboratory (Beltsville, Maryland) is developing a detailed database of reproductive events, including outcomes of veterinary pregnancy examinations, dates of hormonal synchronization, dates of exposure to natural service sires, and designations of “do not breed” for cows that are destined to be culled at the end of lactation.
Genetic Selection for Animal Health
Although genetic evaluation systems are now available for female fertility and longevity, additional improvements are possible. A topic of increasing interest in the United States and Canada is genetic selection for resistance to specific metabolic disorders and infectious diseases. Selection for improved animal health can lead to lower veterinary bills and reduced involuntary culling, as well as enhanced animal welfare. Genetic improvement of animal health will lead to simultaneous improvement of reproductive performance. Using methodology for failure-time analysis, Weigel (2004, unpublished) showed that the relative risk of achieving pregnancy, which was constrained to 1.00 for a healthy, lactating Holstein cow, was reduced by dystocia (0.74), metritis (0.80), retained placenta (0.82), mastitis (0.93), ketosis (0.92), displaced abomasum (0.89), and lameness (0.90), if these events occurred between calving and 75 d postpartum.
At present, only the Scandinavian countries have national data recording and genetic evaluation systems for traits such as clinical mastitis and digestive, locomotive, and reproductive disorders. However, recent research by Zwald et al. (2004a, 2004b) indicated that health data collected from on-farm herd management software programs could be used effectively for genetic selection purposes. Using data from large, commercial farms that used the Dairy Comp 305 (Valley Agriculture Software, Tulare, California), PCDART (Dairy Records Management Systems, Raleigh, North Carolina), or DHI-Plus® (DHI-Provo, Provo, Utah) management software programs, Zwald et al. (2004a, 2004b) grouped farmer-recorded health events into six categories: displaced abomasum, ketosis, mastitis, lameness, cystic ovaries, and metritis/retained placenta. Lactation incidence rates for these disorders ranged from 3% for displaced abomasum to 21% for metritis/retained placenta, and the vast majority of disorders occurred during the first 60 d postpartum. Heritability estimates were 0.14 for displaced abomasum, 0.06 for ketosis, 0.09 for mastitis, 0.04 for lameness, 0.04 for cystic ovaries, and 0.06 for metritis/retained placenta. Predicted probabilities of disease (per lactation) for daughters of individual sires ranged from 1.7 to 6.1% for displaced abomasum, 6.3 to 13.2% for ketosis, 12.9 to 25.9% for mastitis, 7.7 to 13.1% for lameness, 5.9 to 9.1% for cystic ovaries, and 15.1 to 27.1% for metritis/retained placenta. Genetic correlations between disorders were moderate and positive, typically between +0.10 and +0.40; as were correlations between sires’ predicted transmitting abilities for resistance to specific health disorders and their predicted transmitting abilities for DPR. This indicates that genetic selection for improved overall health is possible, as is genetic selection for resistance to specific diseases and disorders.
It is important to note that heterogeneity may exist between farms in the diagnosis, treatment, and recording of specific diseases and disorders. Such heterogeneity can hamper attempts to compare the health status of different herds or to evaluate time trends within specific herds. Because genetic evaluations of dairy sires are based on deviations between the performance of their daughters and the performance of other cows in the same herds at the same time, and because daughters of progeny test sires are distributed across hundreds of herds, small errors or biases that occur within a few herds tend to have a relatively minor impact on estimates of sires’ genetic merit. Nonetheless, validation of the quality and completeness of data from individual farms is critical. Overall, the future appears bright for implementation of selection programs for improved animal health, and corresponding gains in fertility should be expected.
Conclusions
In summary, it is clear that the reproductive biology of today’s high-producing dairy cows differs from that of their ancestors and from that of yearling heifers. High-producing cows tend to have shorter estrous cycles, fewer standing events (of shorter duration), and higher rates of multiple ovulation. Direct genetic selection for improved female fertility is possible using traits such as DPR; and indirect selection, using traits such as longevity or BCS, can provide additional gains. Nearly all leading dairy countries now have systems in place to rank dairy sires based on the reproductive performance of their daughters; but improvements in data collection systems are needed to ensure that information regarding pregnancy examinations, hormonal synchronizations, natural service matings, and do-not-breed designations (of cows to be culled) are routinely reported. Lastly, development of data capture and genetic evaluation systems that can rank dairy sires according to their daughters’ susceptibility to common health disorders, including mastitis, lameness, ketosis, displaced abomasum, and metritis/retained placenta, will enable selection for healthy, long-lasting cows that can achieve pregnancy in a timely manner without excessive management interventions.
Literature Cited
Caraviello, D. Z., K. A. Weigel, M. Florent, A. H. Souza, C. Rawson, N. R. Zwald, and M. L. Wiltbank. 2006. Impact of body condition score on reproductive performance in large commercial dairy herds. J. Dairy Sci. (submitted).
Cartmill, J. A., S. Z. El-Zarkouny, B. A. Hensley, G. C. Lamb, and J. S. Stevenson. 2001. Stage of cycle, incidence, and timing of ovulation, and pregnancy rates in dairy cattle after three timed breeding protocols. J. Dairy Sci. 84:1051-1059.
Cerri, R. L. A., J. E. P. Santos, S. O. Juchem, K. N. Galvão, and R. C. Chebel. 2004. Timed artificial insemination with estradiol cypionate or insemination at estrus in high-producing dairy cows. J. Dairy Sci. 87:3704-3715.
Dechow, C. D., G. W. Rogers, L. Klei, T. J. Lawlor, and P. M. VanRaden. 2004. Body condition scores and dairy form evaluations as indicators of days open in US Holsteins. J. Dairy Sci. 87:3534-3541.
Kadarmideen, H., R. Thompson, and G. Simm. 2000. Linear and threshold model genetic parameters for disease, fertility, and milk production in dairy cattle. Anim. Sci. 71:411-419.
Lopez, H., D. Z. Caraviello, L. D. Satter, P. M. Fricke, and M. C. Wiltbank. 2005. Relationship between level of milk production and multiple ovulations in lactating dairy cows. J. Dairy Sci. 88:2783-2793.
Pryce, J. E., M. P. Coffey, and S. Brotherstone. 2000. The genetic relationship between calving interval, body condition score and linear type and management traits in registered Holsteins. J. Dairy Sci. 83:2662-2671.
Pryce, J. E., B. L. Nielsen, R. F. Veerkamp, and G. Simm. 1999. Genotype and feeding system effects and interactions for health and fertility traits in dairy cattle. Livest. Prod. Sci. 57:193-201.
Royal, M. D., J. E. Pryce, J. A. Woolliams, and A. P. Flint. 2002. The genetic relationship between commencement of luteal activity and calving interval, body condition score, production, and linear type traits in Holstein-Friesian dairy cattle. J. Dairy Sci. 85:3071-3080.
Santos, J. E. P., S. O. Juchem, R. L. A. Cerri, K. N. Galvão, R. C. Chebel, W. W. Thatcher, C. S. Dei, and C. R. Bilby. 2004. Effect of bST and reproductive management on reproductive performance of Holstein dairy cows. J. Dairy Sci. 87:868-881.
VanRaden, P. M., and E.J.H. Klaaskate. 1993. Genetic evaluation of length of productive life including predicted longevity of live cows. J. Dairy Sci. 76:2758-2764.
VanRaden, P. M., A. H. Sanders, M. E. Tooker, R. H. Miller, H. D. Norman, M. T. Kuhn, and G. R. Wiggans. 2004. Development of a national genetic evaluation for cow fertility. J. Dairy Sci. 87:2285-2292.
Zwald, N. R., K. A. Weigel, Y. M. Chang, R. D. Welper, and J. S. Clay. 2004. Genetic selection for health traits using producer-recorded data. I. Incidence rates, heritability estimates, and sire breeding values. J. Dairy Sci. 87:4287-4294.
Zwald, N. R., K. A. Weigel, Y. M. Chang, R. D. Welper, and J. S. Clay. 2004. Genetic selection for health traits using producer-recorded data. II. Genetic correlations, disease probabilities, and relationships with existing traits. J. Dairy Sci. 87:4295-4302.
Author Information
Kent A. Weigel
University of Wisconsin
kweigel@wisc.edu